| <<voltar |
PESQUISA > Peixe elétrico, caramujo e mamíferos > Link 1 > |
Les décharges naturelles de l'organe électrique d'un Mormyre (Gnathonemus petersii) ont une action régulatrice sur le rythme d'autoactivité des électrorécepteurs (Knollenorgan). Ce rythme se groupe très étroitement autour d'une fréquence d'équilibre grâce à l'action physique des activités en bouffées de l'etfecteur. Cette action s'exerce toujours en effet dans un sens qui s'oppose à celui des perturbations qui se produisent pendant l'activité de base.
Les poissons africains de la famille des Mormyridés émettent des décharges électriques (de l'ordre de 10 V hors de l'eau) sous forme de brèves impulsions répétitives qui établissent des champs électriques transitoires dans le milieu aqueux environnant. D'autre part, on peut recueillir, en certains points de leur épiderme, identifiés aux Knollenorgan de Franz, une autoactivité à rythme rapide (d'environ 10 mV hors de l'eau) qui est celle d'un des types d'électrorécepteurs reconnus chez ces poissons [Szabo et Fessard (7), Bennett (3)]. Chaque décharge de l'organe effecteur perturbe le rythme des récepteurs qu'elle atteint. Nous nous sommes demandé si la modification produite est en relation avec le « pattern » des décharges.
L'activité de l'organe électrique d'un Gnathonemus petersii et celle d'un seul électrorécepteur ont été enregistrées simultanément sur bande magnétique dans les conditions décrites par Szabo et Fessard (7) (poisson sous 17 mm d'eau). Les fréquences moyennes de décharge par périodes successives de 1 s (F1) et celles correspondantes de l'électrorécepteur (f1) ont été calculées.
Les séquences de F1, notées F1,i (où I est la date de cette fréquence moyenne), montrent deux types de « patterns » bien distincts :
1. L'activité de base b définie comme toute valeur de F1 à l'exclusion de celles correspondant au « pattern » 2.
2. L'activité en bouffées B, survenant irrégulièrement, et définie arbitrairement par les conditions :
F1,i > et = a 3 imp/s et F1,i + F1,i+1 > et = 5 imp/s.
Les séquences de f1 présentent par contre certaines formes de fluctuations, sans qu'il soit possible de distinguer des types de « patterns ».
L'analyse des séquences temporelles appareillées de F1 (b et B) et f1 a montré que la corrélation du premier ordre est sans signification statistique. Il en est de même lorsque les séquences sont séparément définies, soit par b, soit par B. Cependant, en analysant les séquences de f1 appareillées à F1 on remarque :
1. Une tendance à une plus grande dispersion des valeurs de f1 pendant la durée des activités b (divergence) ;
2. Une tendance inverse, ou convergence, pendant la durée des activités B.
Ce dernier résultat suggère une action de régulation des activités en bouffées (B), ce qui nous a amené à rechercher l'éventualité d'une relation du second ordre entre f1 et les B.
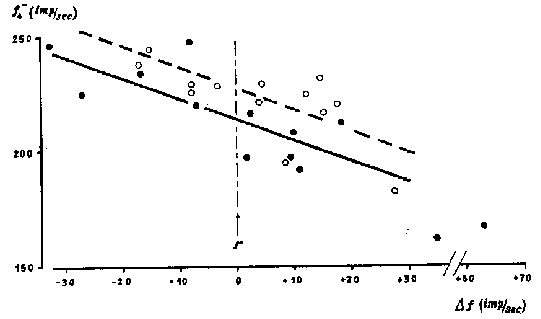
Nous avons calculé la valeur moyenne des f1 pendant les 4 secondes précédant une bouffée, soit f4- et de même pendant les 4 secondes qui la suivent soit f4+.Soit df = f4+.- f4- la différence de ces deux valeurs. La régression linéaire établie pour une série de B montre souvent une corrélation significative entre df et f-. Les coefficients de corrélation peuvent atteindre - 0,90 (avec p < 0,001).
En ordonnées : Fréquence moyenne (f4-) des autoactivités d'un récepteur pendant les 4 secondes qui précédent une bouffée de l'effecteur ; En abscisses : Différence D f entre f4+ (fréquence après une bouffée) et f4- ; Droite solide : Régression avant l'intervention d'un court-circuit (r = - 0,87 et p < 0,001) ; Droite en pointillés : Régression pendant l'intervention d'un court-circuit (r = - 0,71 et p < 0,001).
La droite de régression ( fig.) peut s'écrire :
(1) f4- =k D f +f0
où k est un coefficient négatif et f0 une constante positive.
On peut en conclure que les électrorécepteurs montrent une fréquence moyenne d'activité qui, sous l'action des décharges en bouffées, doivent se rapprocher à long terme d'une fréquence d'équilibre f0. Autrement dit, l'activité momentanément augmentée de l'organe électrique tendrait à régulariser l'activité spontanée des récepteurs.
Pour étayer cette interprétation formelle, nous avons comparé la fréquence moyenne réelle pendant la plus longue période de régulation significative à la fréquence d'équilibre déduite de l'équation précédente. Cette comparaison a été faite dans deux situations expérimentales différentes :
f réelle = 215,64 ± 1,30 imp/s
f0 = 213,4 imp/s.
- Pendant un court-circuit qui introduit une distortion du champ, et pendant 359 s (p < 0,001):
f réelle = 226,91 ± 2,04 imp/s
f0 = 225,4 imp/s.
En tout, 8 électrorécepteurs, dont deux placés dans des conditions physiques différentes, chez cinq poissons dont les décharges présentaient à la fois les deux « patterns » b et B, ont été étudiés selon les mêmes principes. Huit fois sur dix, cette forme de régulation s'est manifestée significativement (p < 0,05). Comme les bouffées constituent l'agent unique pendant les périodes considérées, la cause du phénomène décrit doit être, directement ou indirectement, cet agent.
Il s'agit bien d'une action physique de la décharge et non d'une autre action
contemporaine interne, laquelle n'aurait pas été influencée par la distortion du champ
extérieur.
| <<voltar |
PESQUISA > Peixe elétrico, caramujo e mamíferos > Link 1 > |